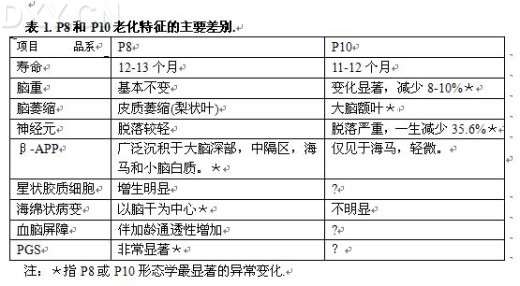
|
快速老化模型小白鼠(Senescence Accelerated Mouse,SAM)由日本京都大学竹田俊男教授经20年近代延交培育而成,具备均一的遗传背景和稳定老化病态特征。 SAM品系中,P8和P10除具有P品系的一般老化症状外,伴快速老化自然出现学习记忆力障碍和低恐怖低紧张状态,为脑老化痴呆模型小白鼠;其共同点是均具有认知障碍,且在渡过生长期后(4~6月),伴快速老化自然发生,与临床痴呆的渐进性发生极为相似。另外两者还各具特点,病理变化具有一定的差异。P8脑中主要出现明显的β-淀粉样蛋白沉积、PAS染色阳性颗粒状结构(PGS)和海绵样病变;P10则伴有广泛性脑萎缩和脑重减轻。其主要差异见表1,是比较理想的研究脑老化和痴呆的模型,与其同源对照组R1相比,其行为学、形态学、神经生化、多种酶类以及蛋白质和基因表达均出现异常,现简述如下。
1、行为学特征 神经行为学实验是研究动物学习记忆力障碍的主要方法。其基本原理是利用动物觅食、求生、逃避伤害、喜暗恶明等本性,设计实验,观察其近期和远期学习记忆能力,评估其痴呆状态。其方法主要有水迷宫、八臂迷宫、明暗箱、穿梭箱、高架桥、跳台等。由于各实验室采取的实验方法不同,在评估学习记忆力障碍发生的时间时报道不一致,但一般均认为,与正常老化的R1相比,P8和P10在8月龄出现明显的学习记忆障碍,处于一种低紧张低恐怖的痴呆状态(P<0.01)。 2、形态学特征 ⑴ 以脑干为中心出现海绵状变化 P8在幼龄时其脑组织即出现海绵状变化,对照组R1小白鼠几乎见不到。如2月龄后以脑干、延髓的网状结构为中心,出现许多空泡性变化并呈海绵状。空泡定量分析表明,空泡的大小和数目伴衰老增加,且与学习记忆缺陷的程度密切相关。用胶质细胞纤维酸性蛋白 (GFAP) 染色,早期可见伴有大量的和广泛的星形细胞增生。另外,空泡性变化在8个月时其峰值下落,至老龄则不明显。 ⑵ 海马锥体细胞的树状棘密度的减少 Golgi染色和组织定量研究表明,P8海马锥体细胞树突棘的密度发生了明显的异常变化,与对照组R1相比,CA1和CA3 棘的密度明显减少,并有统计学意义。说明进入该部位的锥体细胞突触减少。考虑到海马是记忆的重要结构,因此P8的学习记忆障碍与此异常具有直接关系。 ⑶ 随加龄出现明显的星形细胞增生 P8发育早期即出现星形细胞的增生。应用GFAP染色观察,2月龄即可看到斑状的星形细胞增生,推测这可能是对海绵状变化的修复反应。增生发生于多个脑区,最明显的部位与脑干海绵样病变一致。另外,光学显微镜显示海绵状变化不明显的其他部位,如大脑皮质的一部分、纹状体、大脑基底部、脑干背侧等也出现GFAP阳性星形细胞的增加。在上述相应部位,对照组R1只在高龄时观察到显著的星形细胞增生,可能是随加龄的神经细胞脱落、突触变性等多种多样的原因所造成。但是,梨状皮层与其他皮层相比,随加龄的星形细胞增生是比较多的部位,同时出现随加龄皮层厚度变薄的倾向,这种变化像星形细胞增生那样,P8和R1相比有显著差异。 ⑷ 多发PAS(希夫过碘酸)染色阳性颗粒状结构 P8 PAS染色阳性颗粒状结构(PAS-positive granular structure,PGS)在脑内分布最常见的部位是海马、梨状皮层、嗅结节和小脑皮层等,特别是海马的CA1、CA2和CA3最多,除alveus部位之外,几乎遍及全层,并有层状辐射多发的倾向。海马的组织定量研究结果表明,PGS在P8系的3月龄左右可以看到,6月龄后随加龄急速增加。其具体变化过程是,自3月龄开始,PAS阳性颗粒出现于突触后,进一步可见丘脑以及属于边缘系统的梨状区神经核群发生变化。而对照组R1,只出现于少数老龄鼠。P8平均寿命约为R1的一半,即使考虑平均寿命误差,P8 PGS的增加也非常显著。因此PGS的出现及早期多发是其特异点之一。 PGS在老龄鼠的脑内怎样形成仍然是个谜。PGS的特点是一个由数十个颗粒状小体组成的聚集体,用GFA/PAS双重染色,常常在聚集体的中央观察到GFA阳性星形细胞,PGS簇集的大小正好和星形细胞突起延伸一致,一个个小体沿着突起分布。但是也可以考虑这是星形细胞的单纯的反应表现,很可能PGS的形成过程与星形细胞有关。 迄今为止,还没有人和其他动物关于PGS的报告。PGS是SAM新的形态学脑老化指标,同时又在学习记忆障碍的P8多发,这一点很重要。 |
|
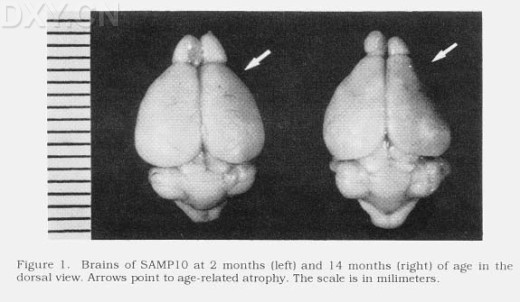
⑸ 血脑屏障的异常 血脑屏障(blood-brain barrier, BBB) 是由毛细血管形成的血浆与脑细胞外液间的屏障,可防止多种外源物质进入脑,但营养物质和代谢产物可顺利通过, 保护脑的内环境高度稳定, 以利于中枢神经系统的机能活动。随着中枢神经系统的老化,BBB的机能低下,导致外源物质易于侵入,出现脑损害。日本学者观察了P8和R1伴加龄血脑屏障的变化,发现血脑屏障的通透性随加龄首先呈现低下,其后则亢进,P8 12月龄与6月龄相比其通透性大约增加了41%(P<0.01), R1 21月龄与12月龄相比增加了26%(P<0.01)。P8与R1比较,P8比R1更早出现异常,12月龄P8比R1大约增加38%(P<0.01)。这些结果说明P8在幼龄即出现初期老化症状,随后伴加龄出现BBB障碍,使大脑更易受到外界物质损伤的影响。 P10是继P8后新开发的脑衰老模型,也以学习记忆缺陷为特点,但具有与P8不同的特征。 ⑴ 显著的脑萎缩 P10伴加龄出现明显的脑萎缩,特别是以大脑皮质前额区为中心,呈进行性发展, 见图1(引自Toshio Takeda所著《The SAM Model of Senescence》), 13-15月龄后,与对照组R1相比脑重减少10%,锥体细胞脱落35.6%。TUNEL研究显示,与学习和记忆密切相关的皮层和海马区的细胞凋亡伴加龄有逐渐增加的趋势,皮层的细胞凋亡,10和8月龄鼠显著高于2月龄及同龄R1; 海马的细胞凋亡,10月龄亦显著高于2、6和8月龄及同龄R1。这与渡过生长期后快速的脑老化相一致,说明细胞凋亡可能是P10脑萎缩的重要原因之一。 图1. SAMP10 2月龄和14月龄脑萎缩状况. 左侧为2月龄鼠脑,右侧为14月龄鼠脑。
⑵ 海马毛细血管伴衰老的变化 Lee等比较了14月龄P10与3月龄R1海马毛细血管的老化状态,发现P10①大多数毛细血管失去光滑轮廓;②毛细血管内腔不规则;③毛细血管内皮增厚;④内皮小囊泡出现更频繁;⑤毛细血管特别是小动脉和静脉的外皮细胞含有许多增大的蜂巢样空泡的胞质包涵体。⑥神经胶质周围血管的盲区增大。这些异常增加了海马血脑屏障功能损伤的可能性和微血管血液动力学紊乱。暗示海马供血异常,可能是其学习记忆障碍的形态基础之一。 3、神经生化特征 神经系统信息的传递以及学习记忆的获得与维持有赖于各种神经递质及其受体适时适量正常地发挥其作用。任何异常都会导致脑功能的障碍和退变。P8所出现的学习、记忆障碍以及低恐怖不安状态,有其形态学基础,也有其生化根据。 ⑴ 葡萄糖代谢的异常 葡萄糖是大脑能量的唯一来源,伴随学习记忆能力的缺陷,葡萄糖代谢出现异常,主要表现为脑内代谢增强,无效利用增加。放射标记葡萄糖发现,14C-2-脱氧葡萄糖在1月龄P8脑中可正常积累,但2-3月龄以后,积累下降,这与P8在2-3月龄开始出现认知损伤相一致。 Omata等通过脑切片研究了P8葡萄糖代谢率和线粒体功能,与R1相比,其2月龄和10月龄葡萄糖的无氧酵解明显增强,而线粒体的功能则下降。6-14C-D-葡萄糖的代谢产物14CO2分析表明,4-8周龄P8大脑细胞悬液14CO2量比同龄R2明显增高,而其组织匀浆液14CO2的量没有差别,说明P8脑中葡萄糖的代谢异常增加,而且这种增加与跨膜的葡萄糖转运系统有关;进一步研究表明脑细胞摄取2-脱氧-D-葡萄糖的能力和细胞松弛素B结合脑细胞膜的能力P8比R2显著增加,而且这种增加只发现于4-8周龄的P8皮层中,在大脑的其他区域和其他周龄鼠(2周龄和40-48周龄)中均没有发现,说明在P8生长发育的早期(1-2月龄)即出现葡萄糖代谢异常和转运蛋白异常,可能此为P8脑损伤原因之一。 ⑵ 一氧化氮的异常 一氧化氮(NO)是一种不同于经典神经递质的信息传递物质,是一种自由基性质的气体。参与血管舒缩状态、免疫功能的调节以及神经信使的传递。可通过强化或消弱突触连接,参与突触调节和突触形成。越来越多的研究证实,NO与动物的学习记忆有关。 最近研究发现,NO与细胞凋亡和神经细胞再生存在一定的相关性。NO和神经元型一氧化氮合酶(nNOS)在年轻和年老的P10皮层中均比同龄R1对照组高,同时检测到抗凋亡蛋白Bcl-2低表达和前凋亡蛋白细胞色素c高表达,进一步采用TUNEL法研究表明,凋亡细胞在P10皮层中更多,说明依赖于衰老的NO增加不仅促进了与Bcl-2相关的细胞凋亡,而且抑制了神经元的的再生,并引起快速脑老化。
自由基对神经细胞的损伤也可能是痴呆的发病主要原因之一。SAMP8、P10正表现为部分脑区神经元退行性变、脑萎缩和脑细胞的变性坏死。Matsugo的研究显示,3,6,9月龄P8脑脂质过氧化物水平均比R1明显增高,且这种增高只与品系有关而与增龄无关,P8在幼龄即出现较高的氧化应激水平是衰老相关的神经退变和损伤的原因之一。 Kurokawa等也研究了P8脑中氧化应激状态,发现10周龄P8大脑皮层的Mn-SOD活性比同龄对照组R1下降50%,但其蛋白含量在两品系中并没有变化。说明Mn-SOD活性的下降与幼龄鼠脑应激反应的增加有关,且Mn-SOD是翻译后修饰而造成的活性降低。 P10也发生了类似的现象。研究显示3、6、9月龄P10脑组织丙二醛(MDA)含量伴加龄明显升高;过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-Px)活性随增龄均呈下降趋势,不同月龄间有显著差异(P<0.05,或P<0.01),与同月龄Rl比较,亦有显著差异(P<0.05,或P<0.01)。说明P8和P10均在幼龄期出现较高的氧化应激水平,一方面自由基增多,另一方面自由基清除酶类的活性显著下降,导致抗氧化防御体系全面下降,在发育早期即可出现生物大分子如脂质、蛋白质、DNA的损伤,导致脑功能障碍。 |
|
⑷ 神经递质及其受体的异常 神经递质在神经系统的信息传递中起着重要作用。研究表明,脑组织老化及伴随的智力低下与神经递质代谢障碍密切相关。据报道AD患者胆碱能系统和去甲肾上腺(NA)系统等存在缺陷,乙酰胆碱(Ach)和乙酰胆酯酶(AchE)减少、NA浓度降低。在P8中,也发现这种异常,中脑、丘脑和视丘下部的Ach较对照组明显下降; 而NA在发育早期其含量就明显低于对照组,而且N-甲基-D-天门冬氨酸(NMDA)受体参与NA的释放并伴增龄降低。免疫组化研究表明P8儿茶酚胺神经元NA的免疫反应在各脑区均比R1明显减弱,免疫电镜显示NA神经元呈现进行性退变。 进一步研究表明,D-丝氨酸与NMDA受体的甘氨酸结合位点结合,增强了谷氨酸与该受体复合体的结合。采用放射自显影技术研究显示,P8海马中D-[3H]丝氨酸与NMDA结合位点的数目较R1明显减少,但其结合位点的亲合性并没有改变。 研究还表明P8还出现氨基酸代谢异常。2月龄和14月龄P8海马和皮层的谷氨酸和谷氨酰胺含量比R1明显升高,高K+诱导的内源性谷氨酸的释放在9月龄和11月龄P8也比R1升高。谷氨酸是一种脑内兴奋性氨基酸(EAAS),其异常增加和蓄积无疑会产生明显的神经毒作用。由于与学习记忆能力密切相关的海马、皮层及纹状体的氨基酸类递质特别是EAAS存在着不同程度的代谢异常,推测这种异常变化导致了神经元坏死,出现相应的学习记忆障碍。 除在P8中发现神经递质及其受体的异常外,P10也有类似发现。与R1相比,P10皮层D2/D3多巴胺受体显著增加,海马则5-HT1A受体显著增加,而在中脑,D2/D3多巴胺和5-HT1A受体均增加,但是D1/D5受体和5-HT2受体却没有什么变化。另外在P10海马中发现,毒蕈碱型乙酰胆碱受体配体结合活性较R1明显降低。 可见P8和P10脑的神经递质及其受体均发生了异常,导致了信息传递的障碍,可能是其学习记忆缺陷等脑功能损伤的重要原因。 ⑸β-淀粉样蛋白积聚 β-淀粉样蛋白沉积是AD重要的病理特征之一,与病情进展和痴呆状态密切相关。Nomura通过免疫组化技术发现P8脑β-APP和27KD的APP片段的免疫反应明显增强。另外,P8脑海绵状周围的星形细胞出现抗APP抗体染色活性,而海绵状病变以脑干为中心广泛发生,说明APP衍生片段在P8脑中显著沉积。 进一步研究发现β-APP及其mRNA在海马中表达增加并与衰老相关;免疫组化分析表明,海马中淀粉样蛋白斑的产生需要APP的表达达到一定的临界量,认知能力的获得与维持的缺陷可通过在脑室中注射β-APP抗体而减轻,说明β-APP的产生与认知能力的缺陷有关。 对8月龄P8海马β-APP进行克隆和序列分析表明,其核苷酸序列与小鼠和大鼠具有99.7%的同源性,与猴子具有88.7%的同源性,与人具有89.2%的同源性;氨基酸序列分析表明,与啮齿类和灵长类分别有99. 2%和97.6%的同源性,其独特之处为第300位的丙氨酸替代了缬氨酸,没有发现类似于AD的突变。APP cDNA能够在Hela细胞中表达并被糖基化,利用特异的APP mRNA反义寡核苷酸可以对其表达时间和表达量进行调控。因此P8的β-APP与AD病人的β-APP极其相似,并有可能通过反义RNA进行治疗。 除在P8脑中发现β-APP沉积外,在P10脑中也发现类似现象。RT-PCR和western blot分析表明,P10海马β-APP蛋白和mRNA 的表达均伴衰老增加,至8月龄达到峰值;而对照组R1的β-APP蛋白水平伴衰老降低,其mRNA水平没有明显的变化。在其他脑区包括皮层、丘脑/中脑和小脑脑干,其蛋白质和mRNA均没有观察到变化。 |
|
⑹ 线粒体功能紊乱 为分析早期线粒体功能紊乱状态,研究了2月龄P8和R1脑线粒体氧化还原状态和氧化磷酸化作用,P8脑线粒体显示了更高的氧化还原状态和更高的线粒体呼吸活性,与R1脑相比,具有较低的呼吸比率,说明一种无效的过量的活动状态在衰老相关的线粒体功能紊乱前就存在于线粒体的电子传递系统。 4、分子水平的研究 衰老的基因学说认为衰老与基因的表达密切相关,是衰老基因按时空顺序顺次表达的结果,快速老化痴呆模型鼠生化指标的异常变化提示,衰老和衰老相关疾病的发生与某些基因和蛋白质的异常表达有关, 因此有学者进一步深入到分子水平探讨其机理。 ⑴蛋白质表达变化 蛋白质是细胞生理功能的执行者,其中有些蛋白如染色质非组蛋白与基因活性的调节密切相关。SDS-PAGE电泳结果显示,3月龄到6月龄P10和R1蛋白质的变化趋势基本一致,但从6月龄到12月龄,62KD的脑细胞核蛋白在P10呈现明显的增加趋势,而R1无明显变化;17KD的脑细胞核蛋白及24KD的染色质非组蛋白在P10呈现明显的下降趋势,而R1仍然持续上升。说明在生长期(3-6月龄)两品系的变化基本一致,生长期到老年期蛋白变化出现明显差异,这与P10的生物学特性,即老化特征的出现在时程上一致。因此其变化可能与SAMP10的快速脑老化密切相关。 神经生长因子(NGF)在前脑胆碱能神经元的维持中起着关键性的作用。免疫组化分析P10 2、5、7、10月龄前脑NGF分布表明,NGF广泛分布于P10的前脑神经元和部分分布于胶质细胞中,并伴增龄减少,推测与P10学习和记忆功能的损伤有关。 Tha等研究发现P8脑中炎性因子也发生了异常变化。2月龄到10月龄,P8和R1两品系脑中白介素(IL)-1β没有明显的差异,但2月龄均比5月龄高,至10月龄则出现差异,IL-1β在P8海马和视丘下部明显增加,其mRNA表达也明显增加;神经坏死因子(TNF-α)和IL-6在大脑皮层和海马升高。说明脑中致炎细胞因子表达的升高可能与P8衰老相关的神经功能紊乱或认知功能缺陷有关。 ⑵ 基因表达的变化 Tobita等采用原位缺口平移法研究了3、6、9和12月龄P8海马区的DNA单链损伤状况,3和6月龄鼠DNA链损伤不明显,与对照组相似;而9、12月龄鼠DNA损伤加重,并伴增龄累加。海马这种伴增龄的损伤很可能与学习和记忆的缺陷有关,而自由基是造成这种DNA损伤的主要损伤原因之一。 为进一步分析衰老对氧化应激基因表达的影响,Kumar等采用了低密度膜芯片研究了4月龄和12月龄P8和C57BL/6鼠海马氧化应激相关基因的变化,发现伴衰老P8 mRNA的表达明显高于C57BL/6,其中T-复合蛋白1eta亚单位在P8中升高了10倍,而C57BL/6只升高了3倍,醌氧化还原酶P8增加了9倍,而C57BL/6只增加了2倍; 另外, 重组活性蛋白(RAG-1)、T-复合蛋白α和β亚单位、细胞色素P450ⅢA25等也较C57BL/6明显增加,说明P8的氧化应激水平显著增加,生物大分子蛋白质、DNA、脂类等更易损伤,造成P8脑功能障碍。 Wei等采用差示法分析了P8海马和R1的mRNA表达差异,发现6个差异表达的基因,序列分析表明W4和W5 cDNA片段属未知基因,W1和W6分别显示出与大鼠eIF-2B和磷脂酶D基因66.1%和62.3%的同源性,W2和W3分别显示与人类大疱性类天疮抗原和人糖原脱分支酶同工酶1/2/3/4/6 89.2%和90.8%的同源性,说明这些基因与P8脑功能障碍有关。 同时,Wei等通过RT-PCR分析了AD相关基因包括β-淀粉样前体蛋白(APP), 早老性蛋白1(PS-1),早老性蛋白2(PS-2),apoE, tau蛋白, c-fos, 神经细胞粘附分子180(NCAM-180), TGF-β1, IL-1α/β, IL-6, TNF-α/β, α-2-巨球蛋白(alpha 2M), II型组织相容性抗原,Bcl-2α,糖皮质激素受体-α(GR alpha)和盐皮质激素受体(MR) 16种基因mRNAs在P8海马和皮层中的表达状况,其中基因TGF-beta 1, IL-1 alpha, TNF-beta, c-fos, NCAM-180, PS-1 和 APP mRNA与R1相比在P8中正常表达,apoE, GR alpha 和 MR mRNAs特别是GR alpha在P8海马中明显低表达,而Bcl-2 alpha, PS-2 和tau蛋白 mRNA则明显高表达;炎性细胞因子(IL-1α/β, IL-6, TNF-β),alpha 2M和MHCII la 抗原没有在SAM脑中检测到。说明P8脑中部分AD基因出现了异常表达,考虑到P8与AD一样具有学习记忆障碍、脑萎缩、β-淀粉样蛋白沉积等特点,因此认为P8具有接近AD的病理特点。 综上所述,P8和P10是一种比较理想的研究脑老化和痴呆的模型,行为学、形态学、神经生化和分子生物学为此提供了较充足的证据。由于它是一种快速自然发病模型,伴快速老化,其脑的形态、结构和功能全面退变,与人衰老和痴呆的临床特征比较接近,不同于造模动物和转基因动物只具有单一的衰老特征。目前,日本、美国、中国、韩国等采用此模型进行了大量衰老相关的基础研究和药物研究,其成果为我们进一步深入理解老化和痴呆的机制提供了基础,愿该模型为更多的研究者认识、了解和应用,促进我国衰老研究的发展。 |
→如果您认为本词条还有待完善,请 编辑词条
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。
0
收藏到: