This is a method for the separation and identification of proteins in a sample by displacement in 2 dimensions oriented at right angles to one another. This allows the sample to separate over a larger area, increasing the resolution of each component.
2D gel electrophoresis is generally used as a component of proteomics and is the step used for the isolation of proteins for further characterisation by mass spectroscopy. In the lab this technique is used for 2 main purposes, firstly for the large scale identification of all proteins in a sample. This is undertaken when the global protein expression of an organism or a tissue is being investigated and is best carried out on model organisms whose genomes have been fully sequenced. In this way the individual proteins can be more readily identified from the mass spectrometry data. The second use of this technique is differential expression, this is when you compare two or more samples to find differences in their protein expression. For instance, you may be looking at drugs resistence in a parasite. In this case you might like to compare a resistent organism to a susceptible one in an attempt to find the changes responsible for the resistence. Here the sequence requirements of the organism are not as important, as you are looking for a relatively small number of differences and so can devote more time to the identification of each protein.
2D electrophoresis is performed in two steps
(1) Isoelectric focusing (IEF) (First dimension electrophoresis): is used in the 1st Dimension (Righetti, P.G., 1983). This separates proteins by their charge (pI).
Isoelctric focusing (IEF) can be described as electrophoresis in a pH gradient set up between a cathode and anode with the cathode at a higher pH than the anode. Because of the amino acids in proteins, they have amphoteric propertites and will be positively charged at pH values below their IpH and negatively charged above. This means that proteins will migrate toward their IpH. Most proteins have a IpH in the range of 5 to 8.5.
Under the influence of the electrical force the pH gradient will be established by the carrier ampholytes, and the protein species migrate and focus (concentrate) at their isoelectric points. The focusing effect of the electrical force is counteracted by diffusion which is directly proportional to the protein concentration gradient in the zone. Eventually, a steady state is established where the electrokinetic transport of protein into the zone is exactly balanced by the diffusion out of the zone. From the factors that regulate the widths of the protein zones and distance between the zones, Svensson and Veterberg derived an equation for the resolution of two similar proteins, based on the following assumptions:
Equation 1: The minimum difference in IpH, for two proteins to be resolved is expressed with equation 1.
From equation 1 it can be seen that by reducing the diffusion, D, the resolution would increase. With a given separation, the only way to accomplish this is to increase the viscosity of the medium. Inert non-charged substances such as sucrose, glycerol etc. may be added or the experiment can be performed in a sieving medium such as a high concentration of polyacrylamide (PAA) gel. Increased viscosity will also affect the mobility (µ ) the mobility of the proteins. This will make the isoelectric separation longer and decrease the resolution by decreasing the dµ /dpH in equation 1_. Therefore, increasing the viscosity is not generally a successful way to improve the resolution although it may explain why there is a clear tendency for better resolution in sieving PAA gels than in more porous agarose gels.
Since the diffusion coefficient is inversely related to molecular size it follows that larger proteins will tend to focus better than smaller ones, other things being equal (See equation 2 _).
The shallower the gradient, dpH/dx (lower values of dpH/dx), the further apart will two proteins be and hence better separated. Note that the factor only applies as the square root. There are some drawbacks with use of extremely shallow gradients: Long focusing times since proteins must migrate a relatively long distance close to the IpH with very low charge: Only the limited number of proteins with IpH values within the narrow pH interval can be analyzed simultaneously; The carrier ampholine may not manage to maintain a completely smooth pH gradient. High field strength (E) will not only increase the resolution, the experimental time is also reduced. Too high field strength may give heat problems if the cooling is inefficient, especially when focusing in the very basic or acid pH region.
The higher the pH dependence of the mobility, (dµ /dpH), the better the focusing. A high electrophoretic mobility close to the IpH will efficiently transfer diffused protein molecule back to IpH. This is essentially an intrinsic factor for the protein that cannot be manipulated. The effect of modifying mobility by affecting the viscosity will be counteracted by the effect of viscosity on diffusion as already discussed and the overall effect is difficult to predict. A high value of dµ /dpH results from the presence of many groups with pKa values close to the IpH. Statistically this is more likely to be the case for a larger than for smaller proteins. Both dµ /dpH and diffusion thus favor the focusing of large proteins and the influence of these factors explains the difficulties of focusing small proteins and peptides to sharp zones.
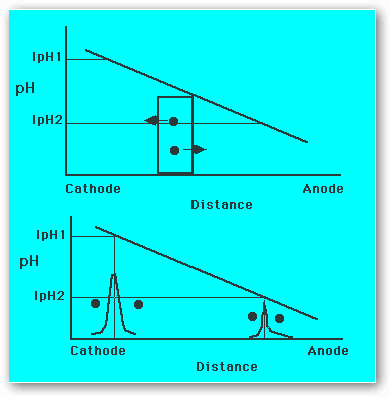
Figure 2: Principle of isoelectric focusing.
Figure 2 is a schematic illustration of a sample with two proteins P1 and P2 place in the center of a pH gradient. P1 is positively charged and will migrate toward the cathode; P2 is negatively charged and will migrate toward the anode. As the proteins approach their IpH, they gradually become less and less charged. The proteins will thus concentrate at the position where pH = IpH. The proteins cannot concentrate in a indefinitely concentrated zone. widening by diffusion is inevitable. Any protein molecule diffusing away from the IpH will acquire a net charge and be transferred back to IpH again by electrophoresis. A balance will be set up between electrophoretic accumulation at IpH and diffusion.
Formation of Natural pH gradients
The formation of a pH gradient is schematically illustrated in Figure 3. Hydrogen ions form at the anode and hydroxyl ions at the cathode in the electrode reactions. This results in region of low and high pH near the anode and cathode respectively and steep pH gradients as one moves into the bulk solution. An amphoteric species with a IpH lower than the average pH in the system will concentrate in the steep gradient close to the anode. A substance with good buffering capacity at its IpH will create a H plateau around its IpH. Given a sufficient number of such substances with evenly distribute IpH values their corresponding plateau will overlap, resulting in a continuous pH gradient. The amphoteric substances that form and stabilize the pH gradient are collectively called carrier ampholytes.
The most essential property for a good carrier ampholyte molecule is a good buffering capacity at its isoelectric point. This requires many pK values close to the buffering capacity at its isoelctric point for each molecular species, making most naturally occurring ampholytic substance, especially most naturally occurring amino acids, useless as carrier ampholytes. Svensson realized that the only way to produce suitable carrier ampholytes was to synthesize substances with the required properties. It was not until the first synthetic carrier ampholytes were successfully prepared that isoelectric focusing could be developed into the practically useful technique of today.
The established pH gradient is maintained by hundreds or thousands of carrier ampholytes molecules lined up in order of IpH with partially overlapping distributions. Since there are no other ionic species in the system, each carrier ampholyte must act as counter ion to other carrier ampholytes consequently each position in the pH gradient will have a unique chemical composition. Electrical conductance and buffer capacity will therefore vary over the pH gradient. Regions with low buffer capacity are more prone to distortion. In preparative experiments with protein loads, buffering capacity form the proteins may affect the pH gradient.
Local heating will occur in the regions with the highest field strength (lowest conductance) and these regions will determine how high an overall voltage can be used. Consequently, other regions with lower field strength will no be focused at optimal conditions. Optimal conditions over the whole pH gradient thus requires even field strength conductance and buffering capacity across the gradient.
A large number of carrier ampholyte mixture are available giving different pH gradients. Many can also be obtained in precast gels ready to use. The optimal pH gradient will depend on the purpose of the experiment. For screening purposes, a broad range interval (pH 3-10 or similar) should be used. A narrow pH range interval is useful for careful IpH determinations or when analyzing proteins with very similar IpH points. Generally, one should not use a narrower gradient than necessary because the shallower gradient will lead to longer focusing times and more diffuse bands. When choosing pH gradient one should be aware that the interval stated by the manufacturer can only be an approximation. The exact gradient obtained depends on many factors such as choice of electrolyte solutions, gradient medium (PAA or agarose), focusing time etc.
Despite the large number of pH intervals available, there may be occasions where none of them fits perfectly. In such cases one can either choose to work with Immobiline or use "pH gradient engineering" in any of the following variants:
::
Extend a given pH interval by adding carrier ampholytes covering the adjacent or a partly overlapping region. Extension into the extreme pH ends can be accomplished by adding acidic or basic compounds.
::
Expand a certain pH area by adding an amphoteric substance, "spacer", such as an amino acid. The spacer should be a "bad" ampholyte so that it does not focus too well.
::
Extend a certain pH range by manipulating the thickness of the gel. The gradient will be shallower in areas with thinner gel. .
::
Manipulating the carrier ampholyte concentration will also affect the steepness of the final gradient. Areas with lower concentration will give shallower gradients.
The different methods can be combined, as was demonstrated by Gill.
Generally IEF will give a true representation of the isoelectric spectrum of the sample. However, IEF of immunoglobulins in standard carrier ampholyte mixtures results in distinct bands in the otherwise continuous smear of immunoglobulin molecules. This was shown to depend on heterogeneity in the carrier ampholyte distribution. For a more truthful representation of the distribution of IpH points in an immunoglobulin sample, the best results were obtained in mixtures of different carrier ampholyte preparations. A mixture of three different Pharmalyte intervals to maximize the number of carrier ampholytes in the interesting region was found to give the best results.
SDS POLYACRYLAMIDE GEL ELECTROPHORESIS (SDS-PAGE) Ed Rybicki and Maud Purves Dept Microbiology University of Cape Town
INTRODUCTION
Electrophoresis is the migration of charged molecules in solution in response to an electric field. Their rate of migration depends on the strength of the field; on the nett charge, size and shape of the molecules and also on the ionic strength, viscosity and temperature of the medium in which the molecules are moving. As an analytical tool, electrophoresis is simple, rapid and highly sensitive. It is used analytically to study the properties of a single charged species, and as a separation technique.
Support Matrices
Generally the sample is run in a support matrix such as paper, cellulose acetate, starch gel, agarose or polyacrylamide gel. The matrix inhibits convective mixing caused by heating and provides a record of the electrophoretic run: at the end of the run, the matrix can be stained and used for scanning, autoradiography or storage.
In addition, the most commonly used support matrices - agarose and polyacrylamide - provide a means of separating molecules by size, in that they are porous gels. A porous gel may act as a sieve by retarding, or in some cases completely obstructing, the movement of large macromolecules while allowing smaller molecules to migrate freely. Because dilute agarose gels are generally more rigid and easy to handle than polyacrylamide of the same concentration, agarose is used to separate larger macromolecules such as nucleic acids, large proteins and protein complexes. Polyacrylamide, which is easy to handle and to make at higher concentrations, is used to separate most proteins and small oligonucleotides that require a small gel pore size for retardation.
Separation of Proteins and Nucleic Acids
Proteins are amphoteric compounds; their nett charge therefore is determined by the pH of the medium in which they are suspended. In a solution with a pH above its isoelectric point, a protein has a nett negative charge and migrates towards the anode in an electrical field. Below its isoelectric point, the protein is positively charged and migrates towards the cathode. The nett charge carried by a protein is in addition independent of its size - ie: the charge carried per unit mass (or length, given proteins and nucleic acids are linear macromolecules) of molecule differs from protein to protein. At a given pH therefore, and under non-denaturing conditions, the electrophoretic separation of proteins is determined by both size and charge of the molecules. Nucleic acids however, remain negative at any pH used for electrophoresis and in addition carry a fixed negative charge per unit length of molecule, provided by the PO4 group of each nucleotide of the the nucleic acid. Electrophoretic separation of nucleic acids therefore is strictly according to size.
(2) SDS- PAGE of proteins (Second dimension electrophoresis)
SDS-PAGE in the 2nd Dimension performs separation of proteins under denaturing conditions This separates proteins by their size (molecular weight, MW). The procedure is known as ISO-DALT: iso for isoelectric focusing and dalt for dalton weight Sodium dodecyl sulphate (SDS) is an anionic detergent which denatures proteins by "wrapping around" the polypeptide backbone - and SDS binds to proteins fairly specifically in a mass ratio of 1.4:1. In so doing, SDS confers a negative charge to the polypeptide in proportion to its length - ie: the denatured polypeptides become "rods" of negative charge cloud with equal charge or charge densities per unit length. It is usually necessary to reduce disulphide bridges in proteins before they adopt the random-coil configuration necessary for separation by size: this is done with 2- mercaptoethanol or dithiothreitol. In denaturing SDS-PAGE separations therefore, migration is determined not by intrinsic electrical charge of the polypeptide, but by molecular weight.
Determination of Molecular Weight
This is done by SDS-PAGE of proteins - or PAGE or agarose gel electrophoresis of nucleic acids - of known molecular weight along with the protein or nucleic acid to be characterised. A linear relationship exists between the logarithm of the molecular weight of an SDS-denatured polypeptide, or native nucleic acid, and its Rf. The Rf is calculated as the ratio of the distance migrated by the molecule to that migrated by a marker dye-front. A simple way of determining relative molecular weight by electrophoresis (Mr) is to plot a standard curve of distance migrated vs. log10MW for known samples, and read off the logMr of the sample after measuring distance migrated on the same gel.
Sample Preparation.
Sample preparation is a key factor in successful 2DE, with complete solubilisation and denaturation of sample proteins being the ultimate aim.
:: Use fresh tissue or sample and do not leave it on the bench. Remember, the final result can only be as good as the starting material. Make sure the sample looks alright, theres nothing better than your eyes for picking out potential problems.
:: Don't handle the sample, always use rubber gloves, clean glassware and equipment. We have found that it is fairly easy to inadvertantly contaminate samples with skin, hair etc from the operator and this is especially important when silver staining is used for protein visualisation. These contaminants, once identified, can act as landmarks, but generally are unsightly and undesirable.
Once again, contaminants will produce unreliable results and will complicate analysis of the completed gel.
Sample Solubilisation.
The preparation of sample for 2D electrophoresis generally requires the use of Solubilisation/Denaturation (SD) buffer. This is used not only to separate the proteins into individual components but also for their denaturation which ensures reliable running in the IEF. Allow time for SD buffer to work, as like many chemical reactions time is required for completion, especially reduction of proteins with DTT. This is best carried out on the bench at room temperature, don't put it on ice, it'll take longer and the urea in the buffer might come out of solution, be aware of potential protease activity though. Hot SDS can be used to increase initial protein solubilisation. However, before electrophoresis it must be removed by competitive displacement in SD buffer. Post solubilisation TCA/acetone extraction may be required (Jefferies et al, 2000) to remove problematic components and this can be combined with SDS solubilisation. If you do use this method be aware that the subsequent protein pellet can be very difficult to resolubilise. We find that the addition of a small volume of 0.1M NaOH can be really useful in overcoming this problem. Start with a few microlitres and increase the volume until the pellet dissolves, agitating the sample as you go, but don't add to much.
Sonication may also help to increase the solubility of the sample, in this case we use a waterbath sonicator.
:: Ultracentrifugation.
This should always be undertaken to clarify the sample before IEF, particulates can cause smearing and may block gel pores. This technique is also very useful for removing unwanted components such as nucleic acids and some polysaccharides.
:: SD buffer components.
The components of the sample buffers we use are listed below, obviously there are other reductants and detergents that can be used for this purpose. :: Chaotrophe: urea and thiourea :: Reductant: DTT, Tris(2-carboxyethyl) phosphine (TCEP), tributyl phosphine (TBP) :: Detergent: CHAPS, triton x-100 :: Ampholytes :: Protease inhibitors
Some researchers also add tris buffer.
Chaotrophe.
Urea is the chaotrope of choice here, its action is through the disruption of hydrogen and hydrophobic bonds, the result of which has no effect on the intrinsic charge of the protein, making it ideal for IEF.
Urea (up to 9M) or urea (7M) & thiourea (up to 2M).
Handling problems: In solution urea forms an equilibrium with cyanate which can cause carbamylation of sample proteins. cyanate production is modulated by pH, temperature and ionic strength.
Precautions:
:: do not heat.
:: buy good quality urea.
:: deionise using ion exchange resins.
:: always use fresh urea solutions or freeze solutions for later use (Means, G.E. & Feeney, R.E., 1971).
Protein carbamylation results in bands of spots across the gel caused by charge change to a particular protein, these are termed carbamylation trains and result from the modification of the peptide amino terminus or the side chains of lysine or arginine by isocyanic acid. Many proteins seem to produce natural trains, when, for instance, they are post-translationarily modified at a number of sites and so exhibit charge heterogeneity. The trick here is to recognise when sample proteins have been modified by carbamylation, a phenomenon that is far from easy to identify. As long as you are careful though, carbamylation should not be a problem.
Thiourea doesn't seem to pose as many problems as urea and is recommended if you are struggling to solubilise your proteins.
Reductants
To fully denature proteins it is necessary to break any disulphide bridges that exist within a particular protein, or that link sub-units of a complex together. This is undertaken by the use of a reductant, of which only relatively mild reducinsg agents are required for conversion of the disulphide bonds of proteins to sulfhydryl groups. There are two major choices:-
Thiols-DTT, beta-mercaptoethanol:
:: For- easy to handle, although Beta-mercaptoethanol is fairly pungent
:: Against- form an equilibrium with reduced disulphides, so higher concentrations are required. DTT also migrates towards the anode during IEF, depleting the basic end of the gel.
Phosphines-TCEP, TBP (Herbert, 1998):
:: For- stoichiometric and only low concs required
:: Against- difficult to handle, TBP spontaniously combusts in the presence of air. TCEP is easier to handle, but is very acidic, you can buy it in a neutral solution but this can add a great deal of salt to the buffer.
Detergents-Surfactants.
Detergents help to disrupt membranes, solubilise lipids and delipidate proteins bound to vesicles or membranes. Hydrophobic interactions can play an important role in the structural integrity of proteins and protein/protein interactions and detergents are ideal for breaking these interactions.
The detergents used in IEF must carry no net charge, so SDS is definitely out of the picture, unless being used for initial solubilisation. They must also be soluble in urea, even at higher concentrations. If they are only sparingly soluble in your buffer any problems may cause them to come out of solution with disasterous consequences. Solubilities vary and as an example we regularly use CHAPS at up to 4% and have no problems, we also use SB 3-10 which is much more problematic, we never use this at higher than 2%.
To be useful the detergent must have strong solubilising characteristics and a great deal of work has been undertaken to find more efficient detergents in this respect (Chevallet et al., 1998). Membrane and strongly hydrophobic proteins can be a real problem and their resolution fuels much interest in this field.
Ampholytes.
The addition of ampholytes to the solubilising buffer (0.5 to 2 % generally) is normally beneficial, aiding protein solubilisation, scavenging cyanate ions and helping in the precipitation of nucleic acids during centrifugation. It has also been suggested that ampholytes are useful in inhibiting interactions between sample proteins and the immobilines of the IPG strips. The ampholyte mixture used should reflect the pH of the IEF gel to be run. For instance, if an pH 3-10 IPG strip is used the ampholyte mix should also be pH 3-10. Ampholytes are stained by both silver and Coomassie and so should be removed from the gel prior to the staining step. Failure to do so may cause a large amount of streaking and smearing at the base of SDS-PAGE gel. We usually fix our gels initially in methanol/acetic acid/water for at least 45 mins to remove them.
Interfering Substances.
a Lipids
b Nucleic acids
c Polysaccharides
d Salts
e Proteases
(a) Lipids.
Not generally a problem for us, except in the case of membrane bound proteins. Lipids can bind to proteins changing both their pI and MW. For this reason it is important to add adequate detergent. Delipidation by solvent extraction (Mastro & Hall, 1999) can be undertaken but may lead to excessive protein loss and may also increase sample variability. In some preparations lipids tend to form a zone at the top of the sample after ultracentrifugation that can be easily removed and discarded.
(b) Proteases.
Can be a severe problem. The denaturing properties of the SD buffer is often sufficient to inhibit the action of many proteases.
There are many types of proteases, so use of the correct protease inhibitors is important. It may be worth checking the literature for information on protease activity in your organism.
Peptide inhibitor cocktails are available that have a wide activity range. These inhibitors are peptides and as such will focus at their pI's, so do not add too much. SDS solubilisation followed by TCA/acetone treatment might improve samples but is not ideal as some proteases have increased activity to proteins denatured with SDS. Careful handling may be crucial, keep the sample cold at all times. But remember, SD buffers with high concentrations of urea should not be chilled too much as precipitation is likely. In our experience the actions and activities of proteases vary greatly between sample types, I have been told by botany people that their proteases are particularly aggressive.
(c) Nucleic acids.
Can block gel pores and increase sample viscosity, they may also bind proteins, particularly nucleic acid binding proteins that will then be depleted and so will not be fully represented on the gel.
Removal: By ultracentrifugation; the higher density of nucleic acids ensures that they are removed without the loss of proteins.
Digest with nucleases; this though brings a number of associated problems. The addition of extraneous proteins, the possible addition of contaminating proteases and an increased complication in the preparation of the sample, we don't bother.
We always ultracentrifuge (30mins at 100 000 x g at 20oC) our samples before we use them anyway and in general find it quick, easy and non-problematic.
(d) Polysaccharides
Uncharged (starch, glycogen) polysaccharides can block gel pores, inhibiting migration of sample proteins resulting in poor focusing. These can simply be removed by ultracentrifugation and should not pose any real problems.
Charged (mucins, dextrans) polysaccharides, on the other hand, bind protein due to charge and so can deplete some proteins. They can also be very difficult to remove.
Polysaccharides cause severe smearing that is evident with both silver and Coomassie staining. we don't have much experience with this problem though.
(e) Salts
High concentrations of salt can pose problems to the integrity of the IEF gel. In our experience it can lead to zones of dehydration in the gel which result in loss of current within the strip. This problem can be especially bad with excretory/secretory products, if the sample has been lyophilised. In this case it is best to dialyse the sample before lyophilisation to remove as much of the salt as possible.
The first IPG strips I ever ran were major disasters due to high salt concentrations. The salts present in the buffer caused the amperage to soar resulting in deformation of the IPG strips which was so severe that the strips fused completely, halting the current . This was a very disturbing experience that I have vowed never to repeat. From this though, I learned 2 valuable lessons, 1) make sure the buffer ion concentration is kept to a minimum. These ions can come from the sample as well as the buffer you use. 2) Always, if at all possible, limit the current to 50mA per strip. If you can't limit the current then keep the voltage down until the amperage in the strips drops. The amperage always starts off high, but as the components in the strip settle down so too will the amperage, which should slowly drop off. Watch the above, you have been warned!
Gel Electrophoresis.
(a) IEF strips.
IPG strips are plastic backed, come in various lengths, but are usually 3.0mm wide and about 0.5mm thick when rehydrated. They come, dehydrated, in a variety of pH ranges. For use they must first be rehydrated, this is generally done under silicone oil. As mentioned earlier it can be beneficial to rehydrate the strips in the presence of your sample (in gel rehydration) and this is the loading method of choice in our lab and enables us to load large quantities of protein. We have used sample cups and found them to be a much less reliable than 'in gel' sample rehydration. We've found there can be problems with sample precipitation at the sample/gel interface and using the sample cups can be awkward and stressful. Other methods have been suggested for applying larger sample volumes (Sabounchi-Schütt et al, 2000) that seem fairly simple to use.
IEF Run.
A number of manufacturers provide equipment for this purpose including Pharmacia, Bio-Rad and others. All this equipment has a number of guidelines which must be adhered to. Temperatures of around 20oC should be used and this should be kept constant as it is a variable and may affect the pI of sample proteins. Increasing the temperature too much above this may result in carbamylation of the proteins. Much lower temperatures may cause precipitation of components such as urea. We find it advantagious to run our strips at low voltage first, then ramp up to higher voltages after the current has settled down. Remember to limit the run by amperage if possible, Bio-Rads Protean IEF is set to 50mA per gel as standard. Optimisation of your particular system will undoubtably be required, however, as there are so many variables you may be required to take some "leaps of faith." A good tip to ensure that the IEF is running correctly is to watch the bromophenol blue front carefully. It should slowly migrate toward the anode. If it does not migrate discretely or breaks down into a number of bands be very suspicious, you may have a problem. Remember though that the bromophenol blue migration cannot be used to measure the progress of the IEF run, it simply gives you a point of reference. Once the run is complete the strips can either be used immediately or frozen down at -70oC for use later.
Strip Equilibration.
The strips must be equilibrated before they can be used for SDS-PAGE. This is necessary for a number of reasons (Gorg et al., 1988) which are reflected in the components of the equilibration buffer. As the second dimension is SDS-PAGE it is important that the proteins are treated with SDS to give them a negative charge. The equilibration step is normally carried out in two steps, both for between 10 and 15 minutes. In the first step DTT is added to ensure that any reformed disulphide bridges are reduced. The second step is the addition of iodoacetamide. This alkylates the proteins and reacts with any unreduced DTT.
Equilibration buffer components:
:: SDS
:: Glycerol
:: Tris, pH 6.8
:: Urea
:: bromophenol blue
To equilibrate the larger 17 or 18cm strips we use plastic pipettes. We cut the pipette end off, place the strip inside, add the buffer, then tape either end up to ensure that it does not leak.
A recent paper by Herbert et al. (2001) suggests that the reduction and alkylation of the sample proteins should be carried out prior to the equilibration step as suggested above. They suggest that the sample should be reduced and alkylated before the IEF step and point out that the pH of the equilibration buffer is far from ideal for the alkylation reaction. They also suggest that reduction without alkylation can disrupt the pH gradient and that due to poor reduction multiple spots can occur through "scrambled" disulphide bridges among unlike polypeptides. Galvani et al. (2001a) provide more information on this and suggests that SDS can strongly quench the alkylation reaction. Galvani et al. (2001b) also found that thiourea in the sample buffer reacts with and quickly depletes the iodoacetamide used for alkylation.
Initially the equilibration suggested by Görg et al. 1988 was to stop point streaking on the second dimension gel, however, this equilibration method has continued and apparently become mainstream. We have always carried out the equilibrations as described and have not noted any major problems. We will, however, now try to reduce and alkylate our samples before any electrophoresis steps to see if there are any improvements.