|
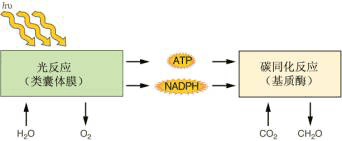
光合作用(photosynthesis)通常是指绿色植物吸收光能,把二氧化碳和水合成有机物,同时释放氧气的过程。地球上一年中通过光合作用约吸收2.0×1011t碳素(6400t/s),合成5×1011t有机物,同时将3.2×1021J的日光能转化为化学能,并释放出5.35×1011t氧气。光合作用是地球上规模最巨大的把太阳能转变为可贮存的化学能的过程,也是规模最巨大的将无机物合成有机物和从水中释放氧气的过程。自从有了光合作用,需氧生物才得以进化和发展。由于光合作用中氧的释放和积累而逐渐形成了大气表面的臭氧(O3)层,O3能吸收阳光中对生物有害的紫外辐射,使生物可从水中到陆地上生活和繁衍。光合作用是生物界获得能量、食物以及氧气的根本途径,所以光合作用被称为“地球上最重要的化学反应”。没有光合作用也就没有繁荣的生物世界。当今人类社会面临着日趋严峻的食物不足、能源危机、资源匮乏和环境恶化等问题,这些问题的解决无一不与植物的光合作用有着密切的关系。因此深入探讨光合作用的规律,揭示光合作用的机理,使之更好地为人类服务,愈加显得重要和迫切。 一、光合作用总反应式的确定 18世纪以前,人们都认为植物是从土壤中获得生长所需的全部元素的。1771年英国化学家普利斯特利(J.Priestley)发现将薄荷枝条和燃烧的蜡烛放在一个密闭的钟罩里,蜡烛不易熄灭;将小鼠与植物放在同一钟罩里,小鼠也不易窒息死亡。因此,他提出植物可以“净化”空气,现在就把1771年定为发现光合作用的年代。以后又经许多人的研究,到了19世纪末,人们写出了如下的光合作用的总反应式: 6CO2+6H2O→ C6H12O6+6O2 (4-1) 应该注意到光合作用反应式中所有的反应物和产物都含有氧,而上面两式并没有指出释放的O2是来自CO2还是H2O。很多年来,人们一直以为光能将CO2分解成O2和C,C与H2O 结合成(C H2O ),然而以下三方面研究证实了光合作用释放的O2来自于H2O 。 1.细菌光合作用 能进行光合作用的细菌称之为光合细菌(photosynthetic bacteria)。光合细菌包括蓝细菌、紫细菌和绿细菌等。其中蓝细菌的光合过程与真核生物相似,紫细菌和绿细菌则不能分解水而需利用有机物或还原的硫化物等作为还原剂。例如:紫色硫细菌(purple-sulfur bacteria)和绿色硫细菌(green-sulfur bacteria)利用H2S为氢供体,在光下同化CO2: CO2+2H2S→(CH2O)+2S+H2O (4-3) 光合细菌在光下同化CO2而没有O2的释放。因此,细菌光合作用是指光合细菌利用光能,以某些无机物或有机物作供氢体,将CO2还原成有机物的过程。 1931年微生物学家尼尔(C.B.Van Niel)将细菌光合作用与绿色植物的光合作用加以比较,提出了以下光合作用的通式: 绿色植物和光合细菌都能利用光能将CO2合成有机物,它们是光养生物。从广义上讲,所谓光合作用,是指光养生物利用光能把CO2合成有机物的过程。 2.希尔反应 1939年英国剑桥大学的希尔(Robert.Hill)发现在分离的叶绿体(实际是被膜破裂的叶绿体)悬浮液中加入适当的电子受体(如草酸铁),照光时可使水分解而释放氧气: 4Fe3++2H2O→4Fe2+ +4H++O2 (4-6) 这个反应称为希尔反应(Hill reaction)。其中的电子受体被称为希尔氧化剂(Hill oxidant),铁氰化钾、草酸铁、多种醌、醛及有机染料都可作为希尔氧化剂。希尔不但证明了给叶绿体照光可使水分解放氧,氧的释放与CO2还原是两个不同的过程,而且也是第一个用离体的叶绿体做试验,把对光合作用的研究深入到细胞器水平,为光合作用研究开创了新的途径。 以后发现生物中重要的氢载体NADP+也可以作为生理性的希尔氧化剂,从而使得希尔反应的生理意义得到了进一步肯定。在完整的叶绿体中NADP+作为从H2O到CO2的中间电子载体,其反应式可写为: 2NADP++2H2O→2NADPH+2H++O2 (4-7 ) CO2也可看作为一种生理性的希尔氧化剂,因为向完整的叶绿体悬浮液中充入CO2或加入能产生CO2的试剂如NaHCO3,照光时叶绿体能发生放氧反应。 3.18O的研究 更为直接的证据是标记同位素的实验。1940年美国科学家鲁宾(S.Ruben)和卡门(M.D.Kamen)等用氧的稳定同位素18O标记H2O或CO2进行光合作用的实验,发现当标记物为H218O时,释放的是18O2,而标记物为C18O2时,在短期内释放的则是O2。这清楚地指出光合作用中释放的O2来自于 H2O。 CO2+2 H218O→(CH2O)+ 18O2+H2O (4-8) 为了把CO2中的氧和H2O中的氧在形式上加以区别,表明光合作用中释放的O2全来自于H2O,而CO2中的一个O又被还原成H2O,因此,可用下式作为光合作用的总反应式。 CO2+2H2O →(C H2O )+ O2+2 H2O (4-9) 二、光反应和暗反应 光合作用需要光,然而是否其中每一步反应过程都需要有光呢?20世纪初英国的布莱克曼(Blackman)、德国的瓦伯格(O.Warburg)等人在研究光强、温度和CO2浓度对光合作用影响时发现,在弱光下增加光强能提高光合速率,但当光强增加到一定值时,再增加光强则不再提高光合速率。这时要提高温度或CO2浓度才能提高光合速率。据测定,在10~30℃的范围内,如果光强和CO2浓度都适宜的话,光合作用的Q10=2~2.5(Q10为温度系数,即温度每增加10℃,反应速度增加的倍数)。按照光化学原理,光化学反应是不受温度影响的,或者说它的Q10接近1;而一般的化学反应则和温度有密切关系,Q10为2~3,这说明光合过程中有化学反应的存在。用藻类进行闪光试验,在光能量相同的前提下,一种用连续照光,另一种用闪光照射,中间隔一定暗期,发现后者光合效率是连续光下的200%~400%。这些实验表明了光合作用可以分为需光的光反应(light reaction)和不需光的暗反应(dark reaction)两个阶段。 1954年美国科学家阿农(D.I.Arnon)等在给叶绿体照光时发现,当向体系中供给无机磷、ADP和NADP时,体系中就会有ATP和NADPH产生。同时发现,只要供给了ATP和NADPH+,即使在黑暗中,叶绿体也可将CO2转变为糖。由于ATP和NADPH是光能转化的产物,具有在黑暗中同化CO2为有机物的能力,所以被称为“同化力”(assimilatory power)。可见,光反应的实质在于产生“同化力”去推动暗反应的进行,而暗反应的实质在于利用“同化力”将无机碳(CO2)转化为有机碳(CH2O)。
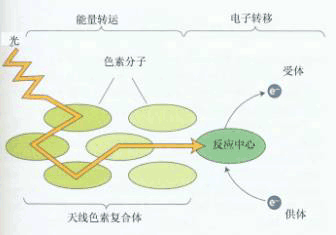
图 4-1 光合作用中“光”反应与“暗”反应的主要产物 光合作用中光反应和碳同化(暗反应)分别在叶绿体的不同区域内。光反应所需要的ATP和NADPH底物合成的一系列反应发生在叶绿体类囊体膜上。光反应产物在碳同化反应中一系列的基质酶的作用下固定CO2转化为碳水化合物。 当然,进一步研究发现光、暗反应对光的需求不是绝对的。即在光反应中有不需光的过程(如电子传递与光合磷酸化),在暗反应中也有需要光调节的酶促反应。现在认为,“光”反应不仅产生“同化力”,而且产生调节“暗”反应中酶活性的调节剂(图4-1),如还原性的铁氧还蛋白。 三、光合单位 释放一个氧分子需要吸收几个光量子?需要多少个叶绿素分子参与?在研究这些问题的过程中,提出了“光合单位”的概念。在研究光能转化效率时,需要知道光合作用中吸收一个光量子所能引起的光合产物量的变化(如放出的氧分子数或固定CO2的分子数),即量子产额(quantum yield)或叫量子效率(quantum efficiency)。量子产额的倒数称为量子需要量(quantum requirement)即释放1分子氧和还原1分子二氧化碳所需吸收的光亮子数。1922年,瓦伯格等计算出最低量子需要量为4,而他的学生爱默生(R.Emersen)等则测定出最低量子需要量为8。后来的实验证据都支持了爱默生的观点,于是8的最低量子需要量得到了普遍的承认,这个数值相当于0.125的量子效率。根据光化学定律(一个分子吸收一个量子,发生一次光化学变化),如果植物的每个叶绿素分子都能进行光化学反应,按还原1个CO2和释放1个O2需吸收8个光量子算,则每当有8个叶绿素分子在一起时,一次足够强的闪光就会造成1个O2的释放。但在1932年,爱默生及阿诺德(W.Arnold)对小球藻(chlorella)悬浮液做闪光试验,计算每次闪光的最高产量是约2 500个叶绿素分子产生1个O2分子,似乎在光合组织中是以2 500个叶绿素分子组成1个集合体进行放氧的,于是当时就把释放1分子氧或同化1分子CO2所需的2 500个叶绿素的分子数目称作1个“光合单位”(photosynthetic unit)。以后又认为,光合是以吸收光量子开始的,应以量子基础计算“光合单位”,1个光合单位应是300(2 500÷8≈300)个叶绿素分子。为什么要300个叶绿素分子吸收1个光子?其解释是:闪光可能被几百个叶绿素分子吸收,可是激发能需传递到1个能够产生光化学反应的“反应中心”(reaction center)区域才能有效。这个反应中心的反应中心色素分子(reaction center pigment)是一种特殊性质的叶绿素a分子,它不仅能捕获光能,还具有光化学活性,能将光能转换成电能。其余的叶绿素分子和辅助色素分子一起称为聚(集)光色素(light harvesting pigment)或天线色素(antenna pigment),它们的作用好象是收音机的“天线”,起着吸收和传递光能的作用。这样就把原来以叶绿素分子数为指标的光合单位看作了能进行光化学反应的光合机构,光合单位成了天线色素系统和反应中心的总称(图4-2)。
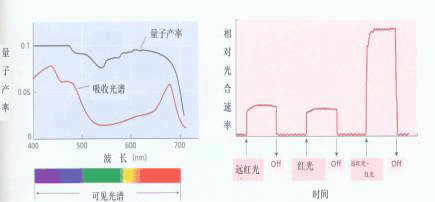
图4-2 光合单位图解 天线色素捕获的一个光量子传递到反应中心色素分子,在那里发生光化学反应进行电荷分离 进一步研究表明,高等植物光反应中电子的传递不只经过一个反应中心,而是要经过两个反应中心,引起两次光化学反应。1986年,霍尔(Hall)等人指出,光合单位应是包括两个反应中心的约600个叶绿素分子(300×2)以及连结这两个反应中心的光合电子传递链。它能独立地捕集光能,导致氧的释放和NADP+的还原。 可见,随着光合研究的深入,“光合单位”的含义已多次被修改。究竟一个“光合单位”包多少个叶绿素分子?这要依据这个“光合单位”所执行的功能而定。就O2的释放和CO2的同化而言,光合单位为2500;就吸收一个光量子而言,光合单位为300;就传递一个电子而言,光合单位为600个叶绿素分子。目前多数人赞同霍尔的看法,认为:所谓的“光合单位”,就是指存在于类囊体膜上能进行完整光反应的最小结构单位。 四、两个光系统 20世纪40年代,以小球藻为材料研究不同光质的量子产额,发现大于680nm的远红光(far-red light)虽然仍被叶绿素吸收,但量子产额急剧下降,这种现象被称为红降现象(red drop)。1957年,爱默生观察到小球藻在用远红光照射时补加一点稍短波长的光(例如650nm的光),则量子产额大增(图4-3),比这两种波长的光单独照射的总和还要高。这种在长波红光之外再加上较短波长的光促进光合效率的现象被称为双光增益效应,或叫爱默生增益效应(Emerson enhancement effect)。以后才知道,这是因为光合作用需要两个光化学反应的协同作用。图4-3中的实线表明≤680nm的光可以对两个光反应起作用,而≥680nm的光只对其中的一个光化学反应起作用。
图4-3 双光增益效应(爱默生效应) 当红光和远红一起照射时光合速率远远大于它们分别照射时光合速率的总和。双光增益效应为位置上的一前一后对波长不同选择的进行光合作用的两个光系统的学说存在提供了有利的证据。 据上述实验结果,希尔(1960)等人提出了双光系统(two photosystem)的概念,把吸收长波光的系统称为光系统Ⅰ(photosystemⅠ,PSⅠ),吸收短波长光的系统称为光系统Ⅱ(photosystemⅡ,PSⅡ)。 另外,从理论上讲一个量子引起一个分子激发,放出一个电子,那么释放一个O2,传递4个电子(2H2O→4H++4e+O2↑) 只需吸收4个量子,而实际测得光合放氧的最低量子需要量为8~12。这也证实了光合作用中电子传递要经过两个光系统,有两次光化学反应。 20世纪60年代以后,人们已能直接从叶绿体中分离出PSⅠ和PSⅡ的色素蛋白复合体颗粒,分析各系统的组成与功能,证明了光系统Ⅰ与NADP+的还原有关,光系统Ⅱ与水的光解、氧的释放有关。 以上仅就光合作用反应式的确定、光暗反应、光合单位、两个光系统等概念的建立介绍了光合作用研究历史中的部分情况。其实,还有许多杰出的成就值得一提。例如,叶绿素分子结构的确定(H.Fischer 1930);光合碳循环的阐明(M.Calvin 1954);叶绿素分子的人工合成(R.B.Woodward 1960);CAM途径的确定(M.Thomas 1960);磷酸化的化学渗透学说的提出(P.Mitchell 1961);叶绿体DNA的分离(R.Sagar M.Ishida 1963);C4途径的确定(M.D.Hatch 1966 C.B.Slack);PSⅡ放氧反应中心复合体的分离(葛培根1982等);光合细菌反应中心三维空间结构的阐明(J.Deisenhofer 1982 H.Michel 1982 R.Huber)光电子传递理论的确定(Marcus 1992);ATP酶的结构与反应机理的研究(Walker 1997 Boyer 1997);……其中有些内容将在下文中提到,有些由于篇幅有限,本章无法安排。 中国的光合作用研究自20世纪50年代开始,取得了长足的进展。如中国科学院上海植物生理研究所在光合作用能量转换、光合碳代谢的酶学研究等方面,中国科学院植物研究所在光合作用的原初反应和光合色素蛋白复合体研究等方面都有所发现和创新。 总之,光合作用研究历史不算长,从1771年至今才200多年,然而由于各国科学工作者的努力探索,已取得了举世瞩目的进展,为指导农业生产提供了充分的理论依据。当前光合作用的研究拟将进一步阐明以下几个关键问题:①光合作用结构与功能的关系及其遗传控制 ②反应中心的结构与功能 ③放氧复合体的结构与功能 ④能量转换与电子、质子传递的规律 ⑤CO2同化调节机理等。只有弄清了光合作用的机理,人类才能更好地利用太阳能,以至模拟光合作用人工合成有机物。此外航天事业的迅猛发展也迫切需要为宇宙飞船、太空空间站乃至为开发其他星球提供氧气和食品等。这些都使光合作用的研究面临新的挑战与机遇。 |
→如果您认为本词条还有待完善,请 编辑词条
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。
0
收藏到: